 Psittacidae
Psittacidae
Il comportamento degli inseparabili
di William R. Dilger
Laboratory of Ornitology
Cornell University
Ithaca, N.Y.
U.S.A.
1962
Tutti gli Inseparabili manifestano lo stretto legame cui si deve a loro antropomorfica denominazione. Le coppie si formano malto presto e generalmente restano unite per tutta la vita. I due partners esteriorizzano il reciproco attaccamento con grande costanza e varietà di modi. Per chi è interessato all'evoluzione dei comportamenti animali gli Inseparabili costituiscono "un campo di ricerca” particolarmente interessante.
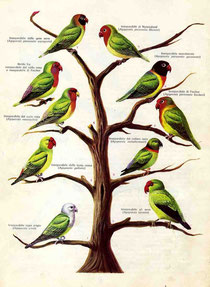
Il Genere comprende nove forme (specie o sottospecie) ben definite che mostrano una diversificazione comportamentale corrispondente alle differenze cromatiche e morfologiche. Attraverso uno studio etologico comparato si può dunque tentare di ricostruire la loro evoluzione e scoprire in che modo la selezione natura1e ha indotto progressi ve modificazioni su uno stesso schema di base.
Con i miei colleghi, nel Laboratorio di Ornito1ogia alla Cornell University, abbiamo studiato negli ultimi cinque anni le costanti e le variabili comportamentali degli Inseparabili. Non è difficile riprodurre in laboratorio le caratteristiche essenziali dell'ambiente naturale africana in modo che chi si possano vivere bene in cattività.
Nel nostro lavoro abbiamo utilizzato tutte le specie di inseparabili ad eccezione di quello a collare nero (Agapornis swinderniana) di cui non siamo riusciti a procurarci alcun esemplare. Le nostre ricerche, in due settori ben definiti: il comportamento sessuale e la costruzione e difesa del nido, si sono rivelate particolarmente fruttuose trattandosi di aspetti dove gli atteggiamenti si sono andati modificando in maniera molto marcata.
Gli inseparabili costituiscono il Genere Agapornis. I loro parenti più prossimi sono i piccoli pappagalli pendenti del genere Loriculus. Tre specie: l'inseparabile del Madagascar, l'inseparabile d’Abissinia e l'inseparabile faccia rossa, somigliano maggiormente ai loricoli e si discostano da tutti gli altri inseparabili per due caratteristiche principali: A) i maschi le femmine hanno una diversa colorazione e sono immediatamente identificati (nelle altre sei specie i due sessi hanno invece identica livrea). B.) il nucleo sociale primario è rappresentato dalla coppia con i figli immaturi (gli altri inseparabili sono fortemente gregari e tendono a nidificare in colonia). Anche da questo punto di vista dunque l'inseparabile del Madagascar, quello da Abissinia e quella faccia rossa, si avvicinano di più alla forma ancestrale.
Il nostro studio sulla diversificazione comportamentale interspecifica ha permesso di chiarire l'ordine cronologico con cui le altre forme comparvero sulla scena. Dopo le tre specie primitive di viene l'inseparabile collare nero, quindi l'inseparabile a collo rossa ed infine le quattro sottospecie di a p, comunemente indicate come inseparabili dall'occhio cerchiato di bianco: l'inseparabile di fischer, l'inseparabile mascherato l'inseparabile del Nassa e l'inseparabile a guance nere.
Vi sono significative differenze comportamentali fra l'inseparabile collo rossa e i quattro dall'occhio cerchiato; ma forse i maggiori contrasti sono quelli che separano le tre specie primitive da quelle che si sono evolute più tardi. Anche la caratteristica comune all'intero genere: la formazione di coppie in giovanissima età, mostrano i due gruppi diversificazione riferibili al modo di nidificare: le singole coppie o in colonia.
Nelle specie primitive Le coppie si costituiscono quando gli uccelli hanno circa quattro mesi di vita. A questa età che si sono completamente indipendenti dai genitori ed hanno già rivestito la livrea adulta. Nelle specie che si sono evolute più di recente, dove la riproduzione in colonia consente la socializzazione tra i coetanei fin dall'abbandono del nido, le coppie si creano ancora prima, quando i pappagalli hanno circa due mesi e rivestono la livrea giovanile.
Tra gli inseparabili la formazione di una coppia è un fatto piuttosto semplice. L'individuo spaiato ne cerca un altro nella stessa condizione che tenta, spulciandolo, di risvegliarne l'interesse. Gli uccelli scoprono rapidamente se siano compatibili e in genere passano poche ore per stabilire un legame che durerà tutta la vita.
Quando le due componenti la coppie raggiungono la maturità sessuale il loro comportamento reciproco di viene molto più elaborato. Questo rituale e, in linea generale, comune a tutti gli inseparabili e alcune attività vengono svolte in modo identico. Altre mostrano viceversa un gradiente delle forme più primitive da quelle che lo sono meno.
Una costante l'atteggiamento indifferente o addirittura aggressivo della femmina nei confronti del maschio che tenta di un approccio. Un'altra è la reazione di quest'ultimo, una combinazione di paura, desiderio sessuale, aggressività e successiva frustrazione. Motivato allo stesso tempo dal timore che desiderio il maschio si avvicina cautamente alla compagna, per allontanarsene poi compiendo una piroetta sul posto Tullio. Questo approccio interrotto lo si osserva in tutte le specie.
Due aspetti del comportamento maschile, inizialmente associati con la frustrazione, mostrano d'altro canto una distinta progressione evolutiva. Il primo è detto “squittio modulato”. Nelle tre specie primitive: l'inseparabile del Madagascar, l'inseparabile da Abissinia e l'inseparabile faccia rossa, il maschio può dire una serie di acute vocalizzazioni ogni volta che la femmina pone termine all'azione di corteggiamento scomparendo all'interno del nido. I suoni variano per intensità e chiarezza e non hanno un ritmo ben definito. Nelle forme di più recente evoluzione: inseparabile collo rossa e quattro dall'occhio cerchiato, le localizzazioni si sono modificate. Il suono ritmico, più puro, di intensità meno variabili e non è prodotto solo quando la femmina volta le spalle e si allontana, ma anche quando è presente e interessata. Lo squittio modulato a dunque subito una progressiva trasformazione non solo per quanto riguarda le caratteristiche fisiche, ma anche per il contesto nel quale si manifesta.
Un'analoga e evoluzione verso un comportamento più fortemente ritualizzato si è verificata in un'altra attività sessuale: il cosiddetto “trattamento sostitutivo", una reazione che deriva dall'abitudine comune tutte le specie, di grattarsi la testa con la zampa quando sono frustrate.
Nelle tre forme primitive il trattamento sostitutivo e ancora vicino alle sue origini. Solo due aspetti lo differenziano da un normale trattamento: il suo contesto e il fatto che venga eseguito sempre con la zampa che si trova dalla parte della femmina. Considerazioni puramente prati che innescano questo comportamento, il fatto cioè che il maschio abbia la zampa già sollevata in previsione dell'accoppiamento.
Nelle specie meno primitive e questo trattamento si trasforma in una manifestazione ritualizzata. Il suo progressivo distacco dalla motivazione originaria appare con sempre maggior chiarezza se esaminato comparativamente nel inseparabile collo rossa e via via nel inseparabile dall'occhio cerchiato di bianco qui il grattamento è molto più rapido approssimativo e non è invariabilmente diretto alla parte piumata del capo. Nel inseparabile collo rossa e allora rivolto al becco, come accade quasi sempre nel inseparabile del Niassa e nell’inseparabile a guance nere. In queste due specie viene usato indifferentemente la zampa dal lato della femmina o l'altra, mentre lo squittio modulato, spesso contemporaneo alla grattamento sostitutivo, è prodotto anche quando la femmina non sembra intenzionata a interrompere il compagno.
In tutte le specie si osserva in mercato di corteggiamento, vale a dire il trasferimento del cibo rigurgitato da un membro della coppia all'altro. Nelle tre specie primitive anche la femmina spesso offre il cibo proprio partner. Questo comportamento non è mai stato osservato nel inseparabile collo rossa e nelle quattro forme dell'occhio cerchiato, dove l’imbeccata sembra una prerogativa esclusivamente maschile.
Si può intravedere uno sviluppo evolutivo nel modo in cui pappagalli seguono il convulso movimento stantuffo del capo, associato con il rigurgito. In a C., a T. ed a P. questi movimenti descrivono un T. collo arco, sono rapidi e numerosi e abitualmente seguiti da contatti fra i becchi piuttosto prolungati, mentre il cibo passa da un individuo all'altro. In a R. N. nelle quattro forme di a P. i movimenti sono più lenti, meno numerosi e tracciano un arco più ampio. Il contatto fra i becchi e generalmente più breve.
Nelle forme che si sono evolute più di recente reazione a stantuffo è andata sempre più assumendo una funzione rituale e non è necessariamente accompagnata dal imbeccata. Le femmine giocano qui in un ruolo ben preciso, mantenendo il piumaggio arruffato durante l'intero processo. Le femmine delle specie primitive non mostrano viceversa alcun particolare comportamento in questa fase.
Le femmine di tutte le specie manifestano la periodica disponibilità al accoppiamento con lievi modificazioni circa il modo di mantenere il piumaggio, soprattutto nel capo. Più la femmina: è il più il maschio è incoraggiato. Alla fine essa solleciterà al accoppiamento piegandosi in avanti sul posatoio e sollevando contemporaneamente la testa e la coda.
Le femmine delle specie primitive, a differenza di quelle che si sono evolute più tardi, non gonfiano le piume durante l'accoppiamento; certamente una conseguenza delle differenze morfologiche fra le varie specie. Poiché nelle forme più recenti entrambi se siano identici colori di segno Le femmine devono rafforzare il riconoscimento da parte dei maschi durante il corteggiamento e l'accoppiamento, con particolari accorgimenti.
Anche se gli atteggiamenti pre-copulatori appaiono innati in tutte le specie, l'apprendimento sembra rivestire un ruolo determinante nei mutamenti che si verificano a mano a mano che due componenti la coppia familiarizzano. Se stesso le coppie di recente formazione sono piuttosto maldestre. In maschi compiono numerosi errori di galateo e sono perciò frequentemente minacciati o interrotti nella loro azione dalle compagne. Dopo qualche nidiata tuttavia, essi acquistano esperienza e tendono sempre più a produrre i giusti comportamenti nei momenti opportuni. Come risultato la femmina reagisce sempre meno aggressivamente e il maschio sempre meno mostra atteggiamenti che implicano frustrazione o corteggiamento interrotto. Anche lo squittio modulato e il grattamento sostitutivo si fanno più rari. Il cauto avvicinamento laterale e ancora praticato, ma con una frequenza sensibilmente inferiore. Nel complesso le manifestazioni pre-copulatorie risultano più brevi. Malgrado l'attività del maschio sia più ridotta la femmina diventa ricettiva piuttosto rapidamente.
I contrasti fra i membri della medesima specie assumono aspetti diversi nel inseparabile di che nidificano per singole coppie rispetto a quelli che lo fanno in colonia. Tra le forme primitive, meno sociali, si è sviluppato un elaborato schema di minacce e pacificazione. Una ben precisa serie di stridii lunghi e ravvicinati, diretti verso un antagonista, segnala ad esempio la volontà di aggredire, mentre il rigonfiamento del piumaggio indica paura e volontà di fuggire. Dopo una serie di reciproche schermaglie il pertinente manifesta la propria sottomissione abbandonando il campo e restandosene quieto, con il piumaggio rigonfio del capo rivolto dalla parte opposta. Per mezzo di questo codice di uccelli possono trasmettere una serie sufficientemente esatta di informazioni circa la propria intenzione di attraccare o fuggire. La conclusione che gli scontri veri e propri raramente si verificano. Quando accadono tuttavia, di uccelli si fanno letteralmente a pezzi.
L'inseparabile collo rossa e le quattro forme dell'occhio cerchiato di bianco, che nidificano colonialmente, entrano in contatto con i membri della propria specie assai più facilmente. Per questo motivo hanno sviluppato una forma di palizzata di combattimento che va ben oltre un semplice codice di aggressione e rappacificamento si e che sostituisce gli scontri fisici. Qui la lotta è soprattutto un duello di becchi, con cui i due uccelli parlano e colpiscono mirando soprattutto alle dita dell'avversario. Le dita sono l'unica parte del corpo che venga realmente beccata. L'inibizione a colpire un rappresentante della propria specie in qualunque altro punto sembra essere, come lo stesso duello colpi di becco, un comportamento innato che l'esperienza può tuttavia perfezionare. La nidificazione in colonia offre ai giovani l'opportunità di fare pratica con i coetanei e di imparare rapidamente.
Se gli inseparabili hanno già avuto occasione di allevare i propri piccoli, essi non alleveranno di quelli di altre specie in possesso di un piumino di colore differente. D'altro canto una femmina la quale, in occasione della prima covata, vengano sostituite le uova con altri specie piumino diverso, alleverà i pulcini che sfileranno, ma rifiuterà nel futuro di occuparsi dei propri figli legittimi. Il piumino del inseparabile collo rossa, come quello delle quattro forme dal occhio cerchiato, e rossiccio mentre negli inseparabili del Madagascar, da Abissinia e a faccia rossa è bianco.
A differenza della maggior parte dei pappagalli, che depongono in una cavità nuda, tutte le specie di Agapornis costituiscono un vero nido. L'inseparabile a faccia rossa colloca il proprio in un buco scavato nel duro fianco del rosso di un termitaio arboreo, mentre gli altri depongono in cavità naturali con un piccolo foro di accesso.
I nidi del inseparabile del Madagascar, del inseparabile da Abissinia e del inseparabile a faccia rossa sono piuttosto rudimentali e consistono in una semplice lettiera sul fondo della cavità.
Queste tre specie hanno elaborato una particolare strategia per la difesa del nido. Nel momento in cui un potenziale predatore farà propria comparsa la femmina gonfia il piumaggio, allarga parzialmente leali e la coda ed emette una sorta di aspro ronzio. Se l'intruso non si allontana essa comprime improvvisamente il piumaggio e con una nota acuta compiono scatto in avanti, senza beccare mandando l'impressione di essere pronta a farlo. Il figlio maggiore può unirsi alla madre in questa manovra, annunciando il piumaggio e facendo udire una specie di brontolio.
L'effetto di questa esibizione è sorprendente e può interrompere l'azione anche di un predatore esperto. Inseparabile del Madagascar, il più primitivo fra gli Agapornis, e il più rapido ad assumere l'atteggiamento di difesa e dell'unico che abbiamo visto portare avanti questa azione attraverso entrambe le fasi. È necessario un forte stimolo perché l'inseparabile da Abissinia si comporti allo stesso modo e non abbiamo mai notato l'uccello andare più in là del semplice rigonfiamento del piumaggio e dell'emissione di aspri suoni raschianti.
Gli inseparabili dal occhio cerchiato di bianco realizzano all'interno della cavità un nido piuttosto elaborato, costituito da una camera d'incubazione cui si accede attraverso una breve galleria. Questa diversa struttura, e la natura fortemente sociale degli uccelli, si combinano rendendo la loro reazione al pericolo diversa da quella delle specie primitive. Non si può parlare di un veri e propri atteggiamento di difesa. Se un predatore raggiunge la cavità il pappagalli interno si limita volare via attraverso il foro d'ingresso (se possibile) o ad appiattirsi sul fondo. Ma se l'intruso incoraggiato da questo atteggiamento di apparente paura, entra nella cavità, si accorge immediatamente che i propri guai sono appena cominciati. Egli infatti deve avventurarsi in una stretta galleria di fissa all'estremità da un avversario dotato di un becco robusto e tagliente.
Va comunque precisato che ha un predatore che raramente concesso di avvicinarsi, perché non appena la sua presenza viene avvertita l'intera colonia assume un particolare atteggiamento intimidatorio. Mantenendo il corpo in posizione verticale degli uccelli battono rapidamente leali ed emettono un verso forte e penetrante. L'effetto visivo sonoro di un intero gruppo impegnato in questa azione è assolutamente straordinario e tale da costituire un valido deterrente per molti potenziali predatori.
Tutte le femmine di inseparabile si procurano ciò che occorre loro per la costruzione del nido più con meno allo stesso modo, con una serie di incisioni parallele in un materiale sufficientemente morbido quale carta, corteccia e fogli. Le mascelle funzionano come una cesoia, tagliando striscioline che variano per forma e dimensioni a seconda della specie.
Gli inseparabili primitivi e inseparabile a collo rosa inseriscono questi frammenti fra le piume del corpo che poi volano via. L'inseparabile del Madagascar, gli inseparabili da Abissinia e l'inseparabile a faccia rossa usano pezzi di materiale di dimensioni molto piccole. Questa è una delle ragioni per cui il loro nido non ha una forma avere propria. Durante la raccolta l'uccello solleva l'intero piumaggio e lo mantiene in questa posizione finché 6-8 frammenti non vi siano stati inseriti. L'inseparabile a collo rossa taglia asserisce sensibilmente più lunghe che consentono la più elaborata struttura del suo nido a cupola. In effetti le strisce sono tanto lunghe da poter essere trasportate solo fra le piume del basso dorso. Questa è la parte del corpo il cui piumaggio viene retto quando il frammenti vi sono infilati. L'inseparabile collo rossa perde circa la metà del proprio carico prima di raggiungere il nido. Alcuni pezzi cadono mentre l'uccello è impegnato a tagliare e inseritile, mentre gli altri si distaccano durante il volo. Gli inseparabili che trasportano frammenti più piccoli sono quelli che dimostrano, da questo punto di vista, una maggiore efficienza.
Questo modo di trasportare materiale da nido esclusivo degli Agapornis e degli affini Loriculus. Qualunque ipotesi sulla loro origine deve partire dalla considerazione che nessun altro pappagallo (con la sola eccezione del Monaco (Myopsitta monachus) sudamericano costruisce un nido per proprio. È quasi certo che questo comportamento ha avuto origine da fattori fortuiti, legati a due caratteristiche attività dei pappagalli: lo spezzettamento di rametti, cortecce e fogli per mantenere il becco affilato e il lisciamento, per conservare un piumaggio pulito e in ordine. Alcuni pappagalli che non costruiscono nidi possono lasciare accidentalmente particelle di sostanze vegetali tra le piume quando interrompono bruscamente il lavoro di frammentazione per passare al lisciamento. Di qui ha avuto molto probabilmente origine l'abitudine di trasportare materiale tra le piume.
Le quattro forme dal occhio cerchiato di bianco si sono totalmente emancipate da questo comportamento ancestrale. L'inseparabile di Fischer, inseparabile mascherato, inseparabile del Niassa e l'inseparabile guance nere, trasportano nel becco materiale da nido, come la maggior parte degli uccelli, con perdite trascurabili durante il tragitto. Impiegano anche rametti, che consentono postazioni più elaborate.
Anche se l'inseparabile collo rossa e solito inserire questo materiale tra le piume, un attento osservatore noterà che una piccola percentuale, un 3% circa, viene portato nel bricco. Questa particolarità ha suggerito un interessante esperimento.
Abbiamo copiato l'inseparabile collo rossa con l'inseparabile di Fischer (questi uccelli s'incrociano facilmente in cattività) per vedere quale comportamento sarebbe emerso da individui geneticamente misti. A sostegno della crisi che il modo di trasportare è sostanzialmente innato, gli ibridi hanno manifestato un conflitto comportamentale fra il metodo dell'inserimento tra le piume (ereditato dal inseparabile collo rossa) e quello del trasferimento nel becco (ereditato dal inseparabile di Fischer).
Quando i nuovi nati cominciarono a fabbricare i primi nidi il loro comportamento denotò un'assoluta confusione. Essi non avevano difficoltà a tagliare le striscioline, ma poi non riuscivano a decidere se portarle fra le piume o col becco. Solo in questo secondo caso (che all'inizio li impegnava per non più del 6% del tempo lavorativo totale) la loro attività era coronata da successo.
Dopo aver ritagliato un pezzetto di foglio di carta tentavano di inserirlo tra le piume del basso d'orzo, ma anche nel caso in cui questa azione aveva successo, il frammento invariabilmente si staccava prima di raggiungere il nido.
Trascorsi circa due mesi, ormai più esperti, gli nidi trasportavano con becco numero di strisce molto maggiore: il 41% per essere esatti. Seguitavano tuttavia compiere movimenti associati con l'inserimento, dirigendo le piume del groppone e del sopra coda e volgendo il capo all'indietro prima di volare via.
Dopo altri due mesi cominciarono a capire che il materiale poteva essere raccolto il trasportato più vantaggiosamente con il becco, solo con un accenno preventivo di inserimento tra le piume. Ma ci vollero due anni perché il seconda atteggiamento diminuisse in modo netto, pur senza scomparire del tutto.
Oggi gli ibridi si comportano più o meno come l'inseparabile di Fischer (di più recente evoluzione rispetto alle specie cui appartiene l'altro genitore) e il solo raramente tentano di inserire strisce nel piumaggio. Sono tuttavia occorsi tre anni per raggiungere questa fase che evidenzia la difficoltà di imparare ad usare un comportamento innato a spese di un altro che pure non è remunerativo. Inoltre, quando seguono i movimenti associati con l'inserimento, si comportano in maniera molto più efficiente di quanto non fosse all'inizio. Evidentemente questa azione non ha bisogno di raggiungere il proprio obiettivo per essere migliorata.
Sino ad oggi i nostri ibridi si sono dimostrati totalmente sterili e dunque non in grado di trasferire il proprio comportamento ad una seconda generazione. Anche nella prima si può tuttavia notare in che modo la natura in 13 elementi nati ed appresi per produrre comportamenti tipici di ciascuna specie.
Ulteriori studi comparativi potrebbe raggiungere molto le nostre conoscenze, non solo per quanto riguarda il comportamento degli inseparabili, ma di che i vertebrati, uomo compreso.
Tratto da:
Bollettino del Centro per lo studio e
la conservazione degli Psittaciformi
N° 3 - 4 1985

